Energy for Health, Vol. 19, Year
A. Colciago1, F. Celotti1, M. Monici2, V. Magnaghi1
1Dept. of Pharmacological and Biomolecular Sciences, University of Milan
2ASA Campus Joint Laboratory, ASA Res. Div & Dept. of Clinical and Experimental Biomedical Sciences “Mario Serio”, University of FlorenceAbstract
An important goal in neuroscience is the study of new therapeutic strategies to promote nerve regeneration in peripheral neuropathies. Evidences from the literature suggest that the application of electromagnetic field (EMF) might be a valid approach to promote nerve regeneration, even if the molecular mechanisms underlying these positive effects are not clearly defined. Aim of our work is to characterize EMF safety and effects on rat Schwann cells (SCs) cultures as an in vitro model of nerve cells involved in axonal integrity. SCs were exposed to EMF with different experimental settings and cell viability, proliferation, migration ability and specific myelin markers were analyzed.
Data suggest that EMF is not a toxic stimulus for SCs even when it is applied repeatedly, moreover a “chronic” EMF exposure induces an increased proliferation without affecting cell differentiation. In conclusion, a repeated EMF exposure might represent a tool to improve regenerative ability of myelin producing SCs on peripheral nerves.
Introduction
Peripheral neuropathies are a group of heterogeneous diseases, characterized by alterations of peripheral nerves structure and functions. Peripheral neuropathies are produced by different etiopathogenetic causes including genetic and metabolic diseases (diabetes, alcoholism, nutritional deficiencies), infective disorders (bacterial or viral), exposure to drugs (e.g. chemotherapy-induced) or environmental toxins and traumatic injuries. Neuropathies are often extremely debilitating and able to significantly compromise the life quality of affected individuals.
The identification of new therapeutic strategies and devices to avoid nerve degeneration and to promote nerve regeneration is therefore an important goal in the field of neuroscience. In literature there are several evidences suggesting the use of electromagnetic fields (EMF) to stimulate peripheral nerves regeneration. Indeed, the application of low frequency fields (20/50 Hz) seems to represent a promising tool to promote nerve regeneration in clinical practice [1, 4]. However, few papers include details regarding the possible cell target affected by the EMF exposure [5, 6]. Therefore, the identification of the molecular mechanisms and the target cells may be fundamental to develop more efficient and strong therapeutic strategies to promote nerve regeneration.
The general aim of our work was to characterize EMF effects and viability on cell potentially related to peripheral nerves regeneration. To this purpose, an in vitro model of rat Schwann cells (SCs), the main peripheral nerve cells responsible of nerve and axonal integrity, were exposed to EMF with different experimental settings.
Materials and Methods
Cell Cultures
Rat SCs cultures were prepared from sciatic nerve accordingly to the method commonly used in the laboratory of Prof. Magnaghi [7]. Briefly, sciatic nerves from 3-day-old rats were digested with collagenase and trypsin, the cell pellets suspended in Dulbecco’s modified Eagle’s medium (DMEM, Serotec, Oxford, UK), supplemented with 10% FCS (Gibco-Life Technologies, Italy) and plated onto Petri dishes. Cells were routinely maintained in DMEM, 10% FCS, 2 μM forskolin, 200 μg/ml bovine pituitary extract (BPE, Invitrogen, Italy); before being used for each different assay, cells were treated for 48h with 4 μM forskolin.
EMF Exposure
According to the specific experimental assays, SCs were plated in Petri dishes and exposed to EMF using the commercial ASA PMT QS device equipped with the Flexa applicator from ASA S.r.l. (Vicenza, Italy). The different protocols and exposure times used are listed in Table I. SCs used as control were plated in same culture conditions, without EMF exposure. For the “chronic” treatment, SCs were exposed 5 consecutive times, every 24 hours each, using the maximum power (3mT), with a frequency of 50Hz for 30 minutes each exposure (Table I).
Proliferation Assay
SCs proliferation was performed by cell count: 60 000 cells were plated into 35 mm petri dishes and collected after 48 and 72 h, with Trypsin 0.05%-EDTA 0.02% in DMEM (PBS, Euroclone, Italy). The cell suspension was then counted under an optic microscope.
MTT Assay
Cell vitality was assessed by MTT (3-(4,5-dimethyl thiazol – 2 – y l )- 2 , 5 – diphenyltetrazolium bromide) assay. This is a rapid colorimetric assay measuring the cell mitochondrial activity. Adherent cells were stained with MTT solution (0.5 mg/ml) for 30 min at 37 °C and absorbance was measured at 570 nm.
Soft Agar
Soft agar colony formation assay is a well-established method for characterizing the anchorage-independent growth of cells in vitro. Cells were grown on a layer of soft agar, thus preventing from adhering to the culture plate. Cells were then exposed to EMF according to the “chronic” protocol (see Table I). After 25 days, colonies were stained with crystal violet and counted.
Scratch Assay
The scratch assay was used to evaluate the migratory ability of SCs. Scratch assay is a reliable and approved method to measure cell migration in response to tissue injury. An artificial gap (so called “scratch”) is experimentally created on a confluent cell monolayer by a mechanical smear. The cells on the edge of the newly created gap move toward the opening, to close the “scratch” until new cell–cell contacts are established again. Images are captured at the beginning and regular intervals during cell migration, and the comparison of the images determines the rate of cell migration. In our experimental conditions, sub-confluent cells in monolayer were scratched with a plastic tip. Wounded monolayers were washed with fresh medium to remove dead cells; images of the scratched monolayers were captured immediately after the scratch (t0), 2h, 6h, 24h and 48h after with the Axiovert 200 microscope (Zeiss, Jena, Germany) and the MetaVue software (Molecular Devices, Sunnyvale, CA, USA). Distances between cell fronts were measured with Image-ProPlus 6.0 (MediaCybernetics, Bethesda, MA, USA), considering at least six measurements from the top to the bottom.
Immunofluorescence
SCs morphology of control and “chronically” EMF exposed cells was evidenced by the cytoskeletal protein actin. Cells were seeded on slides and fixed in 4% paraformaldehyde, then stained with Phalloidin-FITC for f-actin (1: 250, Sigma) and Alexa 488 (green) as secondary antibody (1 : 800, Gibco-Life Technologies). Slides were mounted using Vectashield (Vector Laboratories, Burlingame, CA, USA) and nuclei stained with 4,6-diamidino-2-phenylindole (DAPI). Controls for the specificity of antibodies included a lack of primary antibodies. Confocal microscopy was carried out using a Zeiss LSM 510 System (Gottingen, Germany) and images were processed with Image Pro-Plus 6.0.
Gene Expression
The expression of SCs biomarkers, such as the main myelin-forming protein of the peripheral nerves, glycoprotein P0 and protein PMP22, was assessed by qRTPCR on total RNA.
RNA Extraction, Purification, and quantitation
Total RNA was extracted with trizol (Gibco-Life Technologies) according to the manufacturer’s instructions and then quantified with Nano-Drop2000 (Thermo Scientific, Waltham, MA, USA).
Real-Time PCR
Reverse transcription was performed on 1 μg of total RNA from each sample according to the manufacturer’s protocol (iScript cDNA synthesis kit, BioRad, Segrate, Italy) using random primers. qPCR was done in singleplex in CFX96 Touch™ Real-Time PCR Detection System (BioRad, Segrate, Italy) by using SYBR Green dye (SsoAdvanced SYBR Green Supermix, Bio-Rad, Segrate, Italy) and specific set of primers as follows: P0: 5′-CCTGCTCTTCTCTTCTTTG-3′ and 5′-CACAGCACCATAGACTTC-3′; PMP22: 5′-TCCTGTTCCTTCACATCG-3′ and 5′-TGCCAGAGATCAGTCCTG-3′; α-tubulin: 5′-TCGCGCTGTAAGAAGCAACACC-3′ and 5′-GGAGATACACTCACGCATGGTTGC-3′; β2-microglobulin: 5′-TGCTTGCAGAGTTAAACACGTCAC-3′ and 5′-TTACATGTCTCGGTCCCAGGTG-3′.
Data analysis was performed using the CFX Manager 2.0 software (Bio-Rad, Segrate, Italy). Each sample was analysed in triplicate.
Data was normalized for α-tubulin and β2-microglobulin Ct value. Relative mRNA levels were then calculated by the comparative Ct method (2−ΔΔCt) and data expressed as fold induction.
Statistical Analysis
Data were statistically evaluated by GraphPad Prism 4.00 (San Diego, CA, USA). Statistical significance between groups was determined by means of an unpaired Student’s t-test. P-values < 0.05 were considered as significant.
Results
Figure 1 shows cell viability, measured as cell mitochondrial activity by the colorimetric MTT assay. A single 10 minutes exposure to EMF, applied according to different protocols as described in Table 1 (frequency: 2, 50 and 100 Hz; intensity: 0,15, 1,5 and 3mT), does not induce any statistically significant modification of cell viability.
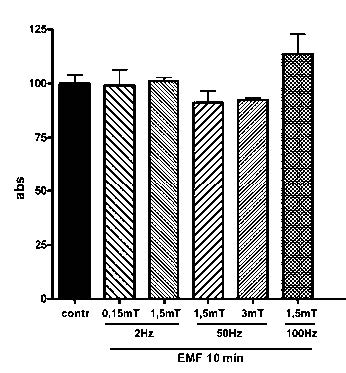
Figure 1
Cell viability, measured as cell mitochondrial activity by the colorimetric MTT assay, in control and EMF-exposed SCs. EMF was applied for 10 minutes with different frequency (2, 50 and 100 Hz) and intensity (0,15, 1,5 and 3 mT). Data are expressed as Absorbance ± SD.
In Figure 2, cell proliferation was evaluated by cell count after EMF exposure (time, frequency and intensity combinations are listed in Table I): EMF was applied once but with different duration (10 and 30 minutes) and cells were counted 48 and 72 hours after EMF exposure. None of the exposure protocols applied produced any significant change in SCs proliferation, neither at low (2Hz) nor at high frequencies (100 Hz), for none of the used intensity (0,15 – 3mT), even when the duration of the exposure was longer (30 minutes).
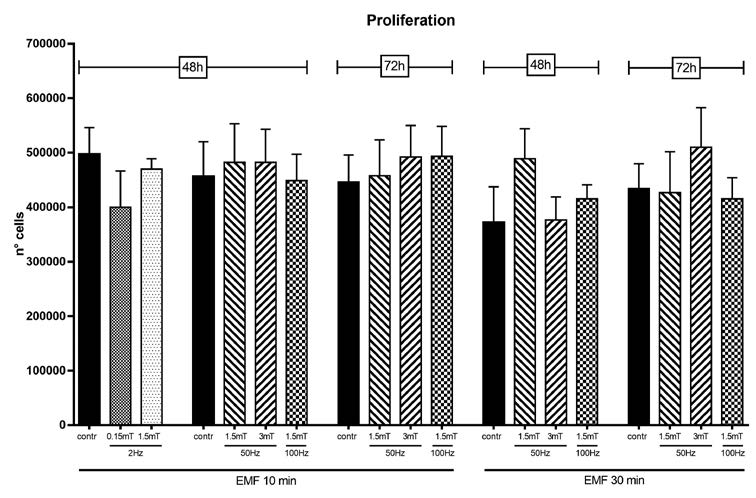
Figure 2
Cell proliferation measured by cell count 48 and 72 hours after EMF exposure, in control and EMF-exposed SCs. EMF was applied for 10 or 30 minutes with different frequency (2, 50 and 100 Hz) and potency (0,15, 1,5 and 3 mT). Data are expressed as number of cells ± SD.
| Frequency | Intensity | Duration | |
|---|---|---|---|
|
2 Hz |
0.15 mT |
(5%) |
10′ |
|
2 Hz |
1.5 mT |
(50%) |
10′ |
|
50 Hz |
1.5 mT |
(50%) |
10′ |
|
50 Hz |
3 mT |
(100%) |
10′ |
|
100 Hz |
1.5 mT |
(50%) |
10′ |
|
|
|||
|
50 Hz |
1.5 mT |
(50%) |
30′ |
|
50 Hz |
3 mT |
(100%) |
30′ |
|
100 Hz |
1.5 mT |
(50%) |
30′ |
|
|
|||
|
50 Hz |
3 mT |
(100%) |
30’ x 5 time (every 24h) |
Table 1
Different combination of EMF frequency and intensity.
Different protocols and exposure times are listed in the Table. Intensity is also indicated as % value relative to max potency of the device used
The scratch assay was used to evaluate the migratory ability of SCs. In a first set of experiments (Figure 3), SCs were exposed for 10 minutes to different combinations of frequency and potency (Table 1), starting from 2 Hz to 100 Hz, the maximum frequency obtained by Flexa. Optical images were acquired at time 0, 2, 6 and 24 hours after EMF exposure; these images were used to measure the distance covered by the migrating front of the cells. Migration ability was calculated as the distance covered in the specific time.
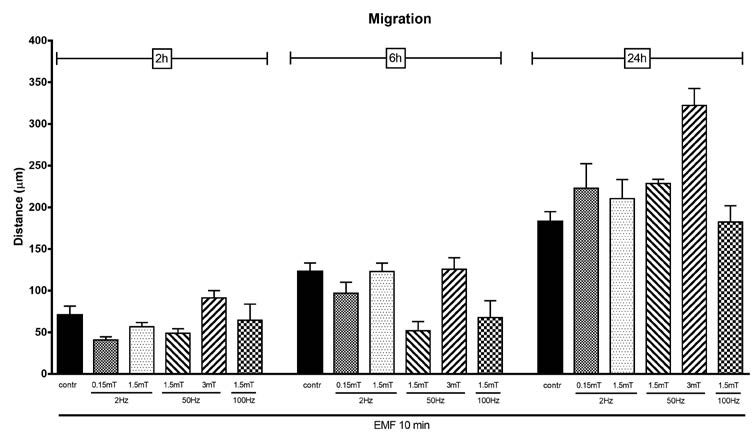
Figure 3
Migration ability evaluated by scratch test in control and EMF-exposed SCs. EMF was applied for 10 minutes with different frequency (2, 50 and 100 Hz) and intensity (0,15, 1,5 and 3 mT). Optical images were acquired at time 0, 2, 6 and 24 hours after EMF exposure; data are expressed as the distance (μm) covered by the migrating front of the cells ± SD.
In a second set of experiments (Figure 4) we focused special attention to the higher frequency and potency, to longer exposure time (30 minutes) and images were acquired up to 48 hours after EMF exposure. As shown in Figure 3 and 4, none of the exposure protocols used is able to significantly modify the migratory capacity of SCs. Cells cover almost the same distance either when are exposed to low (2 Hz) or high EMF frequency (100 Hz). No differences were evidenced at none of the selected EMF intensity (0,15 – 3mT), even when cells were exposed for longer period (30 minutes).
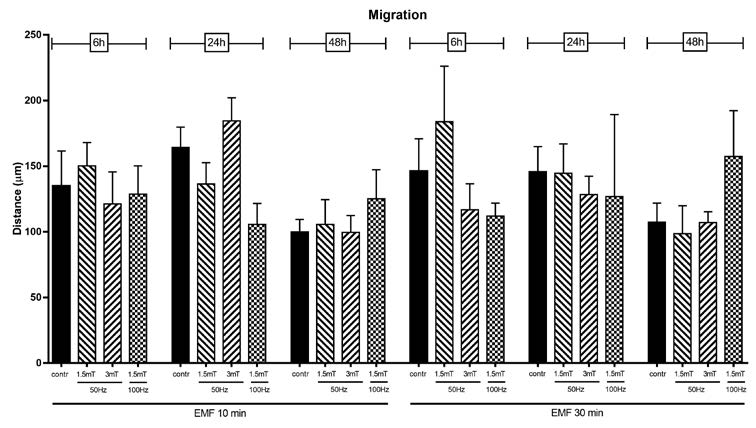
Figure 4
Migration ability evaluated by scratch test in control and EMF-exposed SCs. EMF was applied for 10 or 30 minutes with different frequency (50 and 100 Hz) and intensity (1,5 and 3 mT). Images were acquired 6, 24 and 48 hours after EMF exposure. Data are expressed as the distance (μm) covered by the migrating front of the cells ± SD..
Figure 5 shows a staining of the actin cytoskeleton by phalloidin immunofluorescence in control and EMF exposed SCs. EMF was applied “chronically”, that is 5 consecutive times, every 24 hours each, using the maximum power (3mT), with a frequency of 50Hz for 30 minutes each exposure. Immunofluorescent images show SCs with their classic flat and spindleshaped form, that does not change even after EMF exposure.
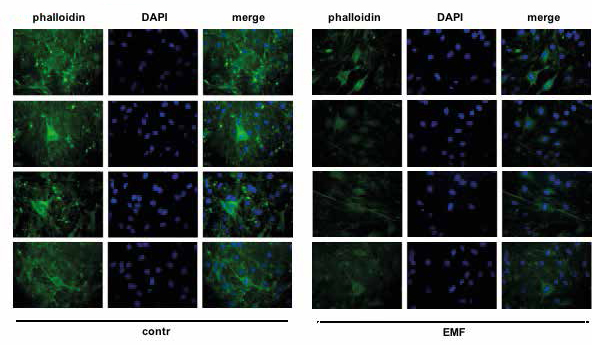
Figure 5
Staining of the actin cytoskeleton by phalloidin immunofluorescence (green) in control and EMF-exposed SCs. Nuclei are stained with DAPI (blue). EMF was applied “chronically”: 5 consecutive exposure every 24 hours each, with a frequency of 50Hz and a intensity of 3mT, 30 minutes each exposure four different fields are shown.
Figure 6 reports the results obtained after the “chronic” EMF exposure as to SCs proliferation, migration and functionality. A prolonged and repeated EMF does not influence either SCs migration rate (Figure 6B), or the expression of P0 and PMP22 (Figure 6D) however, “chronic” EMF exposure induces a statistically significant increase in proliferation (Figure 6A) assessed by cell count, but not in the ability to growth in soft agar (Figure 6C), a measure of the anchorage-independent growth.
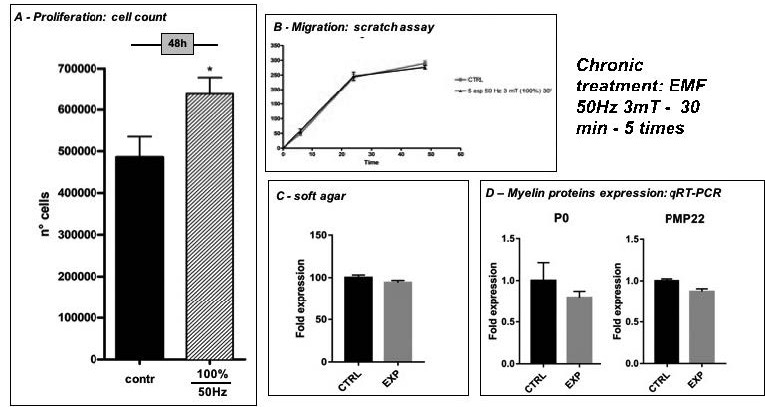
Figure 6
Effects of a “chronic” EMF exposure (5 consecutive exposure -50Hz, 3mT, 30 min – every 24h each) on SCs proliferation, migration and myelin protein expression. A) Cell proliferation measured by cell count 48 hours after EMF exposure in control and EMF-exposed SCs: data are expressed as number of cells ± SD; *p<0,05 vs contr B) Migration ability evaluated by scratch test in control and EMF-exposed SCs: data are expressed as the distance (μm) covered by the migrating front of the cells ± SD. C) Soft agar growth in control and EMF-exposed SCs: the number of stained colonies in EMF-exposed SCs are expressed as a fold induction vs Contr. D) Myelin protein P0 and PMP22 expression assessed by qRT-PCR: data are normalized vs α-tubulin and expressed as fold induction vs control SCs.
Discussion
Cell proliferation and migration are physiologic phenomena strictly related to the regenerative ability of a tissue. This is also true for peripheral nerves and for Schwann cells, the main cell type involved in the regenerative process of nerve tissue. In the data here presented we tested the possibility for EMF to promote nerve regeneration, by means of an “in vitro” approach, using rat SCs as the experimental tool for our studies. Our hypothesis was that if EMF exerts any effect on SCs proliferation and migration, this might be predictive of a regenerative effect of EMF on peripheral nerve. EMF positive effects on nerve regeneration is widely discussed in the literature [4] but there is no general agreement about the application protocol as to the intensity, time intervals and frequency [8]. Also, the molecular mechanisms through which EMF exerts its effects are not well defined, spanning from its influence on NGF levels, cytokines secretion, Ca++ channel modulation, intracellular ROS production etc.; only few of these mechanisms have been also demonstrated in SCs [9]. The different experimental settings used in this paper were designed to clarify some possible mechanisms of EMF action on peripheral SCs. EMF does not represent a toxic stimulus for SCs in culture: the different experimental settings tested, as listed in Table I, never induced any variation in cell proliferation and vitality, even when cells were exposed to EMF with repeated applications. This clearly indicate that SCs maintain an healthy state even when EMF is applied repeatedly and for longer period, thus representing a good experimental model for the evaluation of the effects of EMF chronic exposure on cell proliferation and migration. To our knowledge, this is the first time in which repeated application of EMF is considered as an experimental tool to mimic what happens in clinical use. No positive effect of EMF on cell growth was seen with one short exposure (10 minutes), for none of the different potencies and frequencies applied. Only the EMF repeated exposure for 5 times of 30 minutes each, mimicking a “chronic” in vivo exposure, induces a modest, but statistically significant, increase in SCs proliferation. This is particularly relevant as EMF is applied repeatedly and for a long period of time, when used in therapy.
The cell growth in soft agar, an assay for the assessment of the anchorage-independent growth, was used to evaluate whether the increased proliferation was related to changes towards a less differentiated phenotype. No difference was evidenced between control and EMF-exposed cells, suggesting that SCs increased proliferation does not affect cell phenotype. This is also proved by the expression of the two myelin proteins, P0 and PMP22, considered specific markers of SCs differentiation, the expression of which is almost the same between control and exposed SCs.
In conclusion, our findings evidenced a positive and promising effect of the chronic EMF exposure, generated by the ASA PMT QS, on the SCs in vitro, that may be summarized as follow:
- EMF exposure does not seem to cause toxicity or morphology/differentiation changes on exposed SCs, for 2Hz – 100 Hz frequencies and 0,15 – 3mT intensity.
- SCs morphology, growth, vitality, migration and myelinating capacity are not influenced by low frequency intensity EMF exposure.
- Conversely, a high intensity (3mT), long-time (30 minutes) and repeated (5 times) exposure, even if does not produce signs of cell toxicity, induces an increase in SCs proliferation.
All together, these data are in line with those recently published by our group [10] showing that SCs, exposed to high intensity EMF (50 Hz, 0,1T) for 10 minutes, are able to proliferate and to migrate significantly better than control cells. The effect appears after 24 hours, but it becomes statistically significant for longer exposure times (48 and 72 hours).
Furthermore, a second exposure to EMF, 24 hours later, further increases cell proliferation, suggesting an additive effect. Thus, the EMF, when applied at 50Hz frequency and high intensity (0,1T), exerts a pro-proliferative and pro-migration activity on SCs in culture [10].
We assume that the chronic EMF exposure is promising and might be predictive of regenerative ability following therapeutic application of the device to the peripheral nerves.
Bibliography
- Gordon T, English AW. Strategies to promote peripheral nerve regeneration: electrical stimulation and/or exercise. Eur J Neurosci. 2016; 43(3): 336-50. doi: 10.1111/ejn.13005.
- Al-Majed AA, Neumann CM, Brushart TM, Gordon T. Brief electrical stimulation promotes the speed and accuracy of motor axonal regeneration. J Neurosci. 2000; 20(7): 2602-8.
- Huang J, Zhang Y, Lu L, Hu X, Luo Z. Electrical stimulation accelerates nerve regeneration and functional recovery in delayed peripheral nerve injury in rats. Eur J Neurosci. 2013; 38(12): 3691- 701. doi: 10.1111/ejn.12370.
- Seo NR, Lee SH, Ju KW, Woo JM, Kim BJ, Kim SM, Jahng JW, Lee JH. Low-frequency pulsed electromagnetic field pretreated bone marrow-derived mesenchymal stem cells promote the regeneration of crush-injured rat mental nerve. Neural Regen Res 2018; 13(1):145-153. doi:10.4103/1673-5374.224383.
- Cui M, Ge H, Zhao H, Zou Y, Chen Y, Feng H. Electromagnetic Fields for the Regulation of Neural Stem Cells. Stem Cells Int. 2017; 2017:9898439. doi: 10.1155/2017/9898439
- Galli C, Pedrazzi G, Guizzardi S. The cellular effects of Pulsed Electromagnetic Fields on osteoblasts: A review. Bioelectromagnetics. 2019 May; 40(4):211-233. doi: 10.1002/bem.22187
- Melfi S, Montt Guevara MM, Bonalume V, Ruscica M, Colciago A, Simoncini T, Magnaghi V. Src and phospho-FAK kinases are activated by allopregnanolone promoting Schwann cell motility, morphology and myelination. J Neurochem. 2017; 141(2):165-178. doi: 10.1111/jnc.13951.
- Hei WH, Byun SH, Kim JS, Kim S, Seo YK, Park JC, Kim SM, Jahng JW, Lee JH. Effects of electromagnetic field (PEMF) exposure at different frequency and duration on the peripheral nerve regeneration: in vitro and in vivo study. Int J Neurosci. 2016 Aug;126(8):739-48
- Kerimoğlu G, Güneya C, Ersöz S, Odaci E. A histopathological and biochemical evaluation of oxidative injury in the sciatic nerves of male rats exposed to a continuous 900-megahertz electromagnetic field throughout all periods of adolescence. J Chem Neuroanat. 2018 Sep; 91:1-7.
- Colciago A, Melfi S, Giannotti G, Bonalume V, Ballabio M, Caffino L, Fumagalli F and Magnaghi V. Tumor suppressor Nf2/merlin drives Schwann cell changes following electromagnetic field exposure through Hippo-dependent mechanisms. Cell Death Discovery. 2015; 1, 15021; doi:10.1038/cddiscovery.2015.21


